
Eulimnadia texana are “clam shrimp” that inhabit temporary playas, ditches, and many other ephemeral freshwater habitats throughout the southern United States, west of the Mississippi River, and into northern Mexico (Sassaman, 1989). Hermaphrodites (Fig. 1) produce desiccation-resistant cysts (Fig. 2) which they carry in a brood chamber in the crease of their folded carapace (Fig. 3).

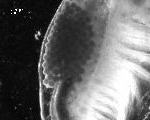
The hermaphrodites bury the eggs within the top several millimeters of the soil. These cysts hatch rapidly following hydration under spring and summer conditions (at water temperatures above 18° C), releasing a nauplius larva (Fig. 4).
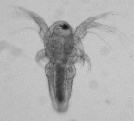
Larval and juvenile growth is extraordinarily rapid. Shrimp reach reproductive size in 4 – 7 days in the laboratory at 27 – 30° C, and in as little as 4 – 6 days in the field. The hermaphrodites produce thousands of eggs in their lifetime, generating clutches ranging between 100-300 eggs, one to two times a day. Hermaphrodites often dig a shallow burrow into which eggs are released. Clutch size increases significantly with carapace length, but no difference in clutch size was detected between selfed and outcrossed clutches in the laboratory.
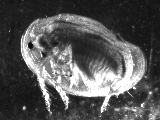
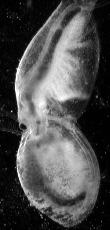
Sexual dimorphism is pronounced. The thoracic appendages of hermaphrodites are unmodified (Fig. 1), but the first two pairs of thoracic appendages in males undergo differentiation into claw-like claspers (Fig. 5) which are used to hold on to the margins of a hermaphrodite’s carapace during mating (Fig. 6).
Eulimnadia texana is omnivorous, able to filter feed as well as forage along pond bottoms.
Natural populations of Eulimnadia are typically hermaphrodite-biased, with some populations completely lacking males. Eulimnadia texana populations range from 0 – 40 percent males, and inbreeding is positively correlated with hermaphrodite-biased sex ratios. Average inbreeding coefficients calculated from six natural populations ranged between 0.20 and 1.0, with an average of 0.49.
Histological examinations of the gonads of E. texana proves the assumption that the populations are composed of males and hermaphrodites. Hermaphrodites have both testicular and ovarian tissue, while males have only testicular tissue (Fig. 7).
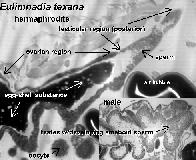
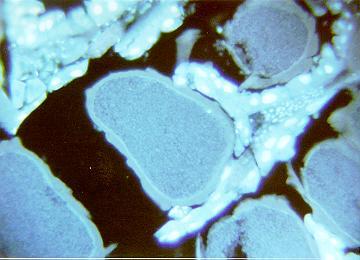
Current histological examinations are aimed at determining when and where fertilization occurs. Developing ova in the ovotestes show no signs of multiple nuclei indicative of fertilization (Fig. 8).
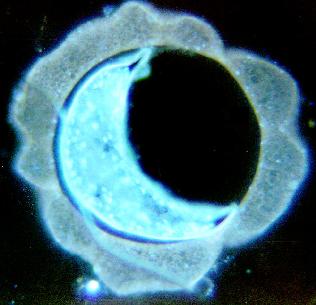
Eggs in the brood chamber show signs of developing embryos (Fig. 9), suggesting that either fertilization is internal but initial cell division does not occur until the eggs are in the brood chamber, or that eggs are actually fertilized externally. Future work is concentrating on delineating these two alternatives.
For further information on the sex determining mechanism of E. texana, check out the article in The American Naturalist